Вопросы экологии применительно к лесному хозяйству
Применительно к лесному фитоценозу гомеостаз характеризуется относительно постоянным числом эдификаторов и упрочением связей между всеми ярусами и видами, входящими в биогеоценоз, вселение «чужаков» в сообщество невозможно, т. к. не имеется свободных ресурсов. Выпадение от модификационных факторов отдельных деревьев верхнего яруса приводит к появлению просвета в пологе, так называемому «ветрова
льно – почвенному комплексу», в виде гантели на месте упавшей кроны и выворота корней. Освободившиеся ресурсы, как правило, заполняются групповым подростом от произрастающих в биогеоценозе видов. Постепенно просветы расширяются до размеров окон. Климаксовый лес всегда неоднороден, в нем имеются многочисленные экологические ниши в виде захламленности, микровозвышений и понижений, разной освещенности, муравьиных куч, кротовин и т. д., что увеличивает биоразнообразие, а значит и устойчивость леса. В целом прирост биомассы климаксового биогеоценоза равен нулю, т. е. в одной части идет нарастание прироста, в другой – падение, в третьей наблюдается равновесие.
Наиболее устойчивы крупные экосистемы, и самая стабильная из них - биосфера, а наиболее неустойчивы – молодые экосистемы. Это объясняется тем, что в больших экосистемах создается саморегулирующий гомеостаз за счет взаимодействия круговоротов веществ и потоков энергии (Ю. Одум, 1975 г.).
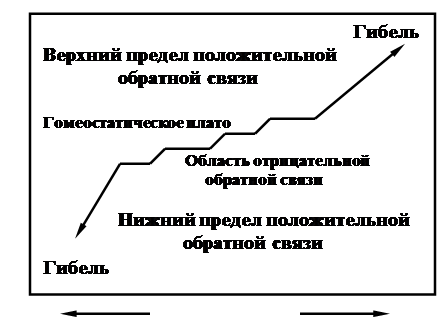
Рис. 6. Представление о гомеостатическом плато, в пределах которого поддерживается относительное постоянство вопреки условиям, вызывающим отклонения.
Вопрос № 28
Концепция климаксового леса: понятие, внутренняя и внешняя структуры, связь с экотопом. Влияние антропогенных факторов на лес.
В предыдущем вопросе при рассмотрении сукцессионных процессов мы уже дали определение климакса, как стабилизированной экосистемы. Первые переселенцы, которые приживаются на новом участке, - это организмы, которые толерантны к абиотическим условиям нового для них местообитания. Не встречая особого сопротивления среды, они чрезвычайно быстро размножаются (саранча, эфемерная растительность и т.п.), т. е. на ранних этапах в эволюции экосистемы преобладает г-стратегия (рост численности). Но постепенно возрастает видовое разнообразие за счет достаточно быстрой смены и увеличения количества популяций и начинает возрастать значение К-фактора (ограничитель роста).
Увеличение видового разнообразия приводит к усложнению связей внутри сообщества, умножению симбиотических связей, снижению чрезмерной рождаемости и доминирования массовых видов, и т. д. Наконец, действия г- и К-факторов уравновешиваются и сообщество развивающейся серии становится стабильным, или климаксным, - «это самоподдерживающееся сообщество, находящееся в равновесии с физическим местообитанием» (Ю. Одум, 1975 г.). Развивающееся сообщество преобразует и само местообитание.
На первых этапах для растительных форм первостепенное значение имеют почвенные биогенные элементы. Но черпать их из запасов почв до бесконечности невозможно и по мере истощения этих запасов разложение отмершей органики становится основным источником питания минеральными веществами биогеохимического круговорота.
Однако такой круговорот возможен лишь в автотрофной системе, черпающей энергию от солнца. Другое дело – гетеротрофная сукцессия, когда приток мертвого органического вещества не восполняет запасы, т. е. первичная продукция равна нулю, и участвуют в сукцессии только гетеротрофные организмы. В этом случае количество энергии не добавляется, а уменьшается, и система прекращает свое существование – все организмы погибают или, в лучшем случае, переходят в покоящиеся стадии. Характерным примером такой сукцессии является сукцессия в гниющих стволах деревьев, в трупах животных, фекалиях и на вторичных стадиях обработки сточных вод. Такая модель сукцессии должна ассоциироваться, по мнению Ю. Одума (1975 г.), с эксплуатацией залежей горючих полезных ископаемых человеком.
На ранних стадиях сукцессионной серии чистой продукции получается значительно больше и при ее изымании человеком сукцессия только приостанавливается, но основа продуктивности на этих этапах не подрывается. Другое дело в климаксных сериях – здесь чистая продуктивность снижается и, в принципе, становится константой. В этом случае очень важно знать величину этой константы с тем, чтобы четко представлять себе ту величину чистой продукции, которую можно изъять из системы, сохранив ее способность к самовосстановлению.
Так, например, вырубку лесов надо вести на локальных участках, с оставлением части территории с коренными типами пород. Это сократит время восстановления фитоценозов, так как сукцессионные серии сократятся до нескольких десятилетий (30 – 50 лет). Чтобы получить полноценную популяцию сосны, состоящую из 30 мозаик от 10-летней биогруппы до 300-летней, необходим период в 300 лет, а чтобы превратить разновозрастную популяцию в одновозрастную рубкой главного пользования достаточно одного года. Сплошная рубка приведет к разрушению всей экосистемы, в том числе ее эдафической части. Это приведет к нарушению сложившейся водонакопительной системы обширного региона, сменит лесной режим, переводящий осадки во внутрипочвенный сток, повлияет на режим открытого места, с резкими колебаниями температур и поверхностным стоком воды. Вырубка переходного типа растительности приводит к появлению пустоши, десятилетиями не зарастающей лесом, а рубка на крутых склонах приводит к потере лесной площади навсегда. Восстановление лишь почв потребует тысячелетия. Более того, сукцессионная серия может пойти и не по пути формирования прежнего лесного сообщества, а, скажем, по пути формирования пустыни и болот или других малопродуктивных экосистем. По теории островной биогеографии (А. В. Яблоков, С. А. Остроумов, 1983 г.), массив леса менее 250 тысяч га не способен обеспечить полноценную жизнь всем лесным видам, неизбежна гибель многих из них из-за недостатка территории для существования минимально необходимого числа особей в популяции.
Таким образом, сообщество не может одновременно быть высокостабильным и давать большой выход чистой продукции, который можно было бы изъять без вреда для самого биоценоза.
В почвенной биоте столь же активно протекают сукцессионные процессы. Они обусловлены разложением органического вещества и лежат в основе биологических круговоротов, - естественных регуляторов процессов, обеспечивающих плодородие почвы. Загрязнение почвенной среды и нарушение процессов образования гумуса снижают регуляторную способность почв и ведут к подрыву естественного плодородия, а следовательно, и к изменениям в экосистеме. Таким образом, эдафическая компонента может весьма существенно повлиять на ход экологической сукцессии при нарушении ее регуляторной функции.
Полнота сукцессий и видовое разнообразие возможны в случае надежной «работы» круговорота питательных веществ. Только в этом случае можно говорить о стабильности экосистемы, которая достигается в результате преобразования сообщества на основе длительной эволюции видов.
Другие рефераты на тему «Экология и охрана природы»:
- Экономические механизмы природопользования
- Расчет сбора за загрязняющую окружающую природную среду
- Киотский протокол - как механизм регулирования глобальных экологических проблем на международном уровне
- Сущность природоохранной функции социально-культурной деятельности
- Экологические законы Барри Коммонера
Поиск рефератов
Последние рефераты раздела
- Влияние Чекмагушевского молочного завода на загрязнение вод реки Чебекей
- Влияние антропогенного фактора на загрязнение реки Ляля
- Киотский протокол - как механизм регулирования глобальных экологических проблем на международном уровне
- Лицензирование природопользования, деятельности в области охраны окружающей среды и обеспечения экологической безопасности
- Мировые тенденции развития ядерной технологии
- Негативные изменения состояния водного бассейна крупного города под влиянием деятельности человека
- Общественная экологическая экспертиза и экологический контроль